

TFD Animal Response
Background
Human geomorphic activity in the United States has re-distributed vast quantities of sediment across the land-scape, particularly in eastern states [Hooke, 1999]. Post-European-settlement deforestation, agriculture, and urbani-zation are responsible for much of the historic landscape changes that have occurred [Trimble, 1974; Costa, 1975; Jacobson and Coleman, 1986; Whitney, 1994]. An important consequence of landscape change has been significant alteration of the distribution, volume, and quality of riverine habitats in response to sediment loading.
Where streams grade into coastal tidal freshwater rivers, historic sediment loadings caused widespread delta progradation and subtidal sedimentation [Gottschalk, 1945; Brush, 1984], and this has fundamentally transformed aquatic communities [Brush and Davis, 1984]. Froomer [1980] analyzed stratigraphic cross-sections of four deltas in Chesapeake Bay tributaries and found that modern alluvium overstepped tidal marshes, completely destroying them. Davis [1985] and Hilgartner [1995] reconstructed paleoecological conditions in upper Chesapeake Bay tribu-taries and found local species extinctions of submerged aquatic vegetation coincident with initial deforestation and land use.
Because many tidal freshwater wetland plants have a competitive advantage in a narrow range of environmental conditions [Pasternack et al., 2000], the initial burial of pre-existing aquatic habitats by watershed sediments was followed by a sequence of allogenic succession driven by continued geomorphic change. At first, unvegetated, frequently flooded mudflats were colonized by floating-leaf plants that survive because of a biological adaptation known as aerenchyma- air spaces in roots and stems which allow diffusion of oxygen from aerial plant parts into roots [Mitsch and Gosselink, 1993]. As river bed elevation rose through time, plant species populations under-went a directional and continuous pattern of colonization and extinction, as demonstrated by paleoecological studies and space-for-time analyses [Khan and Brush, 1994; Hil-gartner, 1995]. A similar process has been reported for saline systems [Ranwell, 1974].
In the face of widespread geomorphic evolution and allogenic plant succession in the tidal freshwater zone, historic wildlife populations must have struggled as their habitats were destroyed. Although it is difficult to determine the pre-settlement population size of animals that lived in the tidal freshwater zone of rivers, records of the early fur-trading industry demonstrate that large numbers of beavers were present on the east coast of the United States at the time of colonization [Whitney, 1994]. Other species such as otters and muskrats were likely to be similarly numerous. Most species have no ability to counteract geomorphic change, but some, notably beaver, may have been able to engineer intertidal and riparian regions to create and maintain preferred habitat conditions. Beavers have long been known to alter riverine habitat through channelization [Berry, 1922], dam building [Ives, 1942; Ruedemann and Schoonmaker, 1938; and Gurnell, 1998], and tree cutting [Jenkins, 1980]. These activities have been reported to have significant ecological and geomorphic consequences, such as development of river floodplains [Ruedemann and Schoonmaker, 1938], increased habitat diversity via patch creation [Johnston and Naiman, 1990; Snodgrass, 1997], and retention of sediment and biogeochemical loads [Naiman et al., 1994; But-ler and Malanson, 1995].
Purpose
Given that fluvial geomorphology, plant ecology, and wildlife populations are all coupled, the question posed in this paper is how did animal populations respond to rapid habitat change in the tidal freshwater zone of rapidly changing tributaries to Chesapeake Bay and did their response effect the interactions between abiotic and biotic variables in the system? Based on the existing literature on beaver activity and the role of beavers in watersheds, it was hypothesized that animals did attempt to counteract habitat changes, so it was necessary to first determine the mechanisms by which such attempts would have been car-ried out in the tidal freshwater zone, as well as the conse-quences of such activity. Mechanisms and animal impacts were investigated by randomized field monitoring at the point scale. Once the current mechanisms by which animals alter habitat conditions were identified and their short-term consequences on biogeomorphology were quantified, a conceptual model for how they would affect geo-morphic evolution over the long term was developed. Actual long-term consequences of observed animal activity at a study site were inferred by interpretation of the present geomorphic and ecological structure of the system in light of the conceptual model.
Study Area
Wildlife observed at OPC include Castor canadensis (beavers), Ondatra zibethicus (muskrats), and Lontra canadensis (river otters). Signs of animal activity, includ-ing debarked trees, channel bank slides, animal lodges, and channelized marsh plains are widespread on the delta. No ecological studies of these species or others have been conducted at OPC.
Methods
Otter Point Creek was under study 1990-1998 to better understand the interactions between physical and ecological processes in the tidal freshwater tributaries of upper Chesapeake Bay. Even though a study of animal activities was not initially planned, the extent of activity combined with the random siting of monitoring transects enabled sufficient observation to warrant this investigation. Mapping and monitoring of vegetation, sedimentation and erosion, substrate conditions, and animal activity were carried out April 1995 to November 1997 at OPC. During an 88-week period from April 1995 to March 1997, monitoring focused on the small HaHa Branch Wetland (HBW) on the north flank of OPC where the HaHa Branch stream terminates. In the middle of that study, monitoring was expanded to sites throughout OPC that were observed for a 60-week period from September 1996 to November 1997. In all, 53 monitoring sites were used, and these were distributed along randomly located transects, as detailed by Pasternack and Brush [1998] and Pasternack [1998].
Data Analyses
Monitoring sites impacted by animal activity were com-pared with non-impacted sites in the same plant association to distinguish the biotic disturbance signal from back-ground abiotic processes. One set of comparisons focused on time series of net sedimentation rates and percent or-ganic content of deposited material in relation to animal activity at individual sites. A second set of comparisons focused on time-averaged process-morphology feedbacks among all study sites as evident in relations between sediment response variables (for example, sedimentation rate) and biogeomorphic controls (for example, habitat index and elevation). All analyses were combined to obtain a conceptual model of the impacts of animal activity on bi-weekly to interannual time scales.
Once the conceptual model was developed, implications of animal activity for delta evolution over longer time scales were assessed. Indicators of the actual long-term impacts of animal activity were identified by studying the structure of delta landforms as well as by considering distributions of plant and wildlife habitats. Broad scale spa-tial patterns were investigated using field observations coupled with Geographical Information System (GIS) data. GIS data included a 1.22 m resolution 1994 color infrared digital orthophoto of the delta as well as plant association, topographic, and sedimentation pattern data layers.
Results
Monitored Animal Activities
Four out of the fifty-three monitoring sites were directly impacted by animal activity. The animals responsible for the impacts were beavers and muskrats, with each causing a different effect and working during a different time of the year. Both animal species uprooted plants, mixed the surface sediment layer, dug new marsh plain channels, maintained old marsh plain channels, made mud piles, built new lodges, and maintained old lodges. Whereas beaver impacts were initially rapid and spanned a large radius, muskrats continuously worked over time and focused cutting along their channels. Also, beaver activity occurred in autumn while muskrat activity occurred in spring and early summer.

In HBW, beavers also cut trees in the riparian forest on the eastern side of the marsh and used those trees in an attempt to dam the HaHa Branch stream just upstream of an abandoned paleo-distributary channel. The riparian forest that was the source of the wood for the dam had 313 trees, which were fortuitously all mapped, identi-fied, and measured for diameter at breast height six months before dam building commenced. Arboreal species found in the forest included Prunus serotina (black cherry), Acer rubrum (red maple), Liquidambar styraciflua (sweet gum), Ilex opaca (American holly), Betula nigra (river birch), Salix nigra (black willow), and Prunus virginiana (choke cherry). Acer rubrum was the most prevalent species, and it was the primary one used for the dam. This population was very young, with diameters ranging from 2-4 cm in most cases.
The dam failed during spring high flows in 1996 and was never rebuilt. Instead, beavers used the remaining debris to make a large lodge in the middle marsh habitat on the western side of the channel adjacent to the paleo-channel.

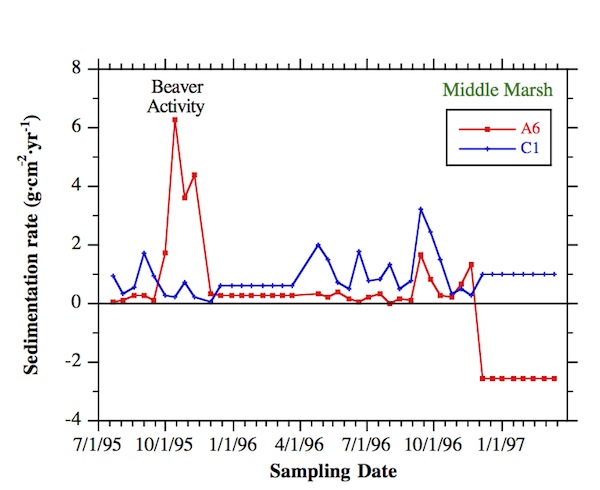
Site A6 was in an HBW area containing middle marsh and some high marsh plants that were all herbaceous. Beavers altered it in autumn 1995 by cutting down most plants in a 180-m2 region and re-excavating a channel net-work that was barely visible prior to their effort due to sed-iment deposition during the previous year. Where they passed through the high marsh and middle marsh near A6, the rehabilitated beaver channels were  rectangular and 30 cm wide x 20 cm deep, with dredged sedi-ment piled on top of channel banks. The main stem of the excavated network that ran through the low marsh was not rectangular because the sediment was not cohesive enough to sustain straight-sided banks, and there were no plant roots on the banks to aid cohesion. Field observations dur-ing biweekly sampling trips indicated that the ground sur-face elevation in the impacted area was decreased relative to the surrounding land, though no repeat elevation surveys were possible. Associated with the decreased elevation was a clearly visible, increased flooding depth and duration relative to the surrounding undisturbed area.
rectangular and 30 cm wide x 20 cm deep, with dredged sedi-ment piled on top of channel banks. The main stem of the excavated network that ran through the low marsh was not rectangular because the sediment was not cohesive enough to sustain straight-sided banks, and there were no plant roots on the banks to aid cohesion. Field observations dur-ing biweekly sampling trips indicated that the ground sur-face elevation in the impacted area was decreased relative to the surrounding land, though no repeat elevation surveys were possible. Associated with the decreased elevation was a clearly visible, increased flooding depth and duration relative to the surrounding undisturbed area.
According to monitoring data from site A6, beaver activity locally increased sedimentation for a 2-month period. This increase was a direct result of lowered elevation and increased flooding, which enabled more sedi-ment transport. No measurable variation in organic content of accumulating sediment was observed at A6 during the beaver activity, because organic content was low in these sediments and organic matter has very low mass. Nevertheless, the sediment was quite distinguishable in appearance, looking like muddy hay, with many fragments of shredded plant stalks and roots. Sedimentation rates at A6 dropped down to background values in 1996, though vegetation underwent reverse allogenic suc-cession to low marsh species, particularly Peltandra virginica (arrow arum), providing further evidence for lower elevations and increased flooding. However, in the mild winter of 1997 the site was significantly eroded as were other low marsh sites reported by Pasternack and Brush [1998] and unlike other middle marsh sites, illustrating long term consequences of animal disturbance. Pulses of organic deposition occurred in autumn 1996, and were caused by foliage dropped from the adjacent riparian forest buffer zone.
A comparison of time series among middle marsh sites shows that the signal of beaver disturbance at A6 is signif-icantly different from the background abiotic signal. Sites C1 and A7 are the other middle marsh sites monitored at HBW. Because winter 1997 erosion at A6 counterbal-anced the earlier deposition, the mean net sedimentation of the three sites is similar. However, the standard deviation in sedimentation was much higher at A6 than at the other sites. Neither A7 nor C1 showed a significant spike in sedimentation in October 1995, demonstrating the local nature of the disturbance, even though A7 was only 30 m away from A6. Toward the end of the beaver activity, some sediment was carried further into the marsh where it deposited at site A7, causing the only significant temporal variation in sedimentation rate at that site over the whole study. In terms of organic content, A7 showed consistently higher values- twice those of the other two sites- due primarily to its inability to flush autumn arboreal debris from the fringing, forested buffer zone except in early spring when no vegetation was present on the marsh surface. Beaver activity in the vicini-ty of A6 caused so much surface sediment mixing in autumn 1995 that arboreal debris could not accumulate there. Also, in the fall of 1996, the beaver-created channel near A6 was still effective enough to continue to export arboreal debris. No other site along the A-transect was able to ex-port accumulating debris, so regardless of which mechanism- surface mixing or channel export- was more im-portant, beaver activity was an important factor in the fate of a major source of organic deposition.
Site B5 was in a Phragmites australis (common reed) patch in the high marsh, and this patch occurs on the edge of a sand splay caused by overbank flooding of the HaHa Branch stream. An actively used muskrat lodge was present at the site throughout the study. B5 showed a high average sedimentation rate and a low average organic content. A spike in organic content at B5 in May 1996 was an artifact of extremely low sedimentation. Under this condition, accumulating material was predomi-nantly composed of in situ plant leaves whose deposition is capricious. Significant over-winter deposition and frequent large variations in sedimentation rate distinguished the muskrat-impacted site from comparable sites without the influence. For example, B6 was a high marsh site 30 m further upslope in the marsh that was not in the stand of reeds. That site showed little temporal variation in sedimentation rate except for a single spike of inorganic sedimentation in the first half of April 1996. The cause of the spike at B6 was unknown.
 Site 2-P15 was in a middle marsh area dominated by Typha angustifolia (cattails). In late summer and autumn 1996, beavers cut down all cattails in a 40 m2 area near 2-P15 and used them to build a lodge. As at A6, the impacted area was visibly depressed and was flooded for a greater duration. The result of this change was a high average sediment-deposition rate and a low average organic content. Sedimentation rates in late summer and autumn 1996 were moderate, while organic contents were low. On December 25, 1996 a dead beaver was found on the western side of the cleared area. During winter 1997, the area around the lodge, including site 2-P15, experienced significant net erosion, which was typical of low marsh habitat rather than the middle marsh habitat that was present prior to beaver activity. During the following spring, muskrats took over the abandoned lodge and dug small channels throughout the area. These muskrat channels connected the site to the marsh front 10 m away and a major distributary channel 25 m to the east. Meanwhile, few new middle marsh plants sprout-ed in the impacted zone, though they grew in the adjacent area. Instead, the low-marsh plant Peltandra virginica colonized the disturbed ground. Since this plant decom-poses quickly upon death, it is unsuitable for constructing lodges. Consequently, very little material was available for lodge maintenance, and the lodge was observed to dete-riorate rapidly during summer 1997.
Site 2-P15 was in a middle marsh area dominated by Typha angustifolia (cattails). In late summer and autumn 1996, beavers cut down all cattails in a 40 m2 area near 2-P15 and used them to build a lodge. As at A6, the impacted area was visibly depressed and was flooded for a greater duration. The result of this change was a high average sediment-deposition rate and a low average organic content. Sedimentation rates in late summer and autumn 1996 were moderate, while organic contents were low. On December 25, 1996 a dead beaver was found on the western side of the cleared area. During winter 1997, the area around the lodge, including site 2-P15, experienced significant net erosion, which was typical of low marsh habitat rather than the middle marsh habitat that was present prior to beaver activity. During the following spring, muskrats took over the abandoned lodge and dug small channels throughout the area. These muskrat channels connected the site to the marsh front 10 m away and a major distributary channel 25 m to the east. Meanwhile, few new middle marsh plants sprout-ed in the impacted zone, though they grew in the adjacent area. Instead, the low-marsh plant Peltandra virginica colonized the disturbed ground. Since this plant decom-poses quickly upon death, it is unsuitable for constructing lodges. Consequently, very little material was available for lodge maintenance, and the lodge was observed to dete-riorate rapidly during summer 1997.
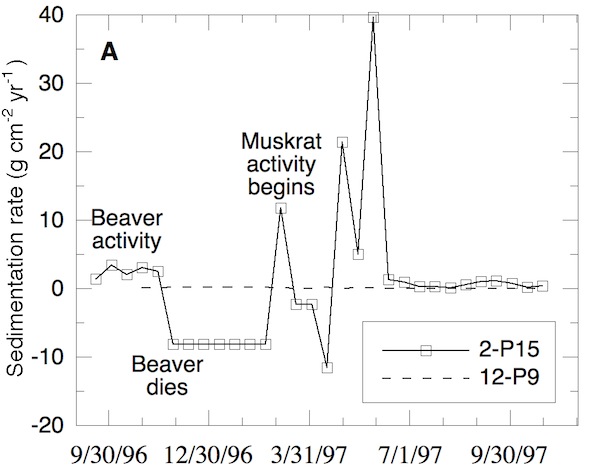
Concomitant with the muskrat activity, extreme swings in sedimentation rate followed. The peak sedi-mentation rate was 40 g·cm-2·yr-1 while the minimum was 0.12 g·cm-2·yr-1. Unfortunately, the data cannot distinguish whether the high loads and fluctuations were due to changes in the rate of sediment import or changes in local redistribution, but both mechanisms are directly caused by animals in this instance. By comparison, non-animal impacted sites such as 12-P9 had almost no measurable deposi-tion or erosion throughout the study. Under background conditions, OPC middle marsh areas receive less than 0.07 g·cm-2·yr-1. Site 2-P15 experienced more than an order of magnitude more sedimentation than that on average.
Site 4-P3 was in a middle marsh locale with a mixture of arrow arum and cattails. Surrounding the disturbed area was a densely populated cattail field. The monitoring site showed many signs of animal activity in recent years, including in-filled animal channels, cut cattail stalks, and a dilapidated lodge. Muskrats began working the site in spring 1997. During the following summer, they intensively channelized the area and built up a former beaver lodge with stalks from surrounding cattails. In contrast to beaver activity observed at A6 and 2-P15, muskrats did not graze all vegetation indiscriminately, just plants where channels would go. As a result, the site developed pockets of arrow arum mixed in with remaining cattail stands. Sedimentation and erosion dynamics at 4-P3 reflect the timing of muskrat activity. During autumn for both years, the sedimentation rate was a moderate 1-2 g·cm-2·yr-1. During winter 1997, the site experienced substantial erosion, while during summer 1997 it experienced a large pulse of deposition. The average sedimentation rate over the study was the highest at the disturbed site, at 1.18 g·cm-2·yr-1, and had the highest standard deviation. Meanwhile, organic content in the disturbed area was the lowest, though there was a large pulse of or-ganics in autumn 1997 when cattail debris was deposited.
Animals in Marsh Ecogeomorphology
When sedimentation rates are plotted as a function of habitat index, disturbed sites show a predictable process-morphology feedback in relation to the delta’s biogeomorphic evolution line. The autumn 1995 time-averaged sedimentation rates for non-impacted HBW sites show a strong response to habitat index. The relation suggests that biogeomorphic parameters such as elevation and plant associations control sedimentation processes at the seasonal time scale. As discussed by Pasternack et al. [2000], there is a dynamic interdependence between abiotic and biotic processes, such that plant associations and ground elevation are in turn changed by sedimentation over annual to interannual time scales. When a space-for-time transformation is applied to the seasonal relation, it appears that as a site evolves into higher and drier habitats, sedimentation rates decrease exponentially under normal conditions.
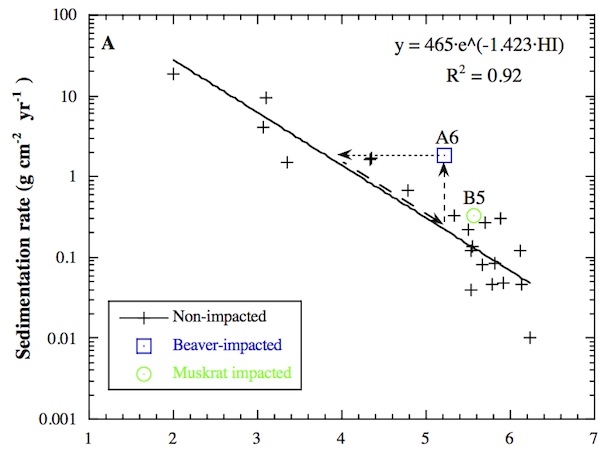
In contrast with non-impacted sites, the beaver-impacted site (A6) plotted far off the trend line during the time of beaver activity, while the muskrat-impacted site (B5) showed only a small deviation. Arrows in the above figure graphically represent site A6’s evolution. The decreased elevation and increased inundation induced by beavers at A6 caused an immediate increase in sedimentation (vertical arrow) followed by a reverse succession in the plant association toward low marsh plants during the next growing season (horizontal arrow), as described earlier. Thus, the data from A6 shows that when plant species and sedimentation rates are not mutually adjusted due to a rapid disturbance, there is a natural process response that returns the system to the equilibrium balance between abiotic and biotic variables defined by the line in the figure. Lower elevation, more flooding, and higher localized sedimentation requires a shift to more inundation- and sedimentation-tolerant species.
Once equilibrium is re-established, the higher sedimentation rate of the new regime causes elevation to increase faster, thereby undoing the animals’ work. Ultimately the site should return to its pre-disturbance habitat state over time (diagonal arrow), requiring animals to periodically maintain their channels and cleared areas. Overall, the system has a negative feedback mechanism that counter-acts local animal activity. Because biweekly sedimentation data is not suitable for making accurate long term calculations, it is not possible to accurately estimate how long it would take the site to revert back to middle marsh, assuming that animals allow such a reversion to occur. Most likely this process takes several years, depending on how much winter erosion occurs under the imposed low marsh regime.
Whole Delta-Scale Animals Impacts
The general geomorphic structure of the OPC delta is typical of shallow, semi-enclosed, fluvial-dominated re-ceiving basins. Beyond the easily identifiable forested, intertidal, and subtidal zones, there are characteris-tic channel features such as multiple distributary channels, abandoned channels, and “middle ground” bars where channels end. Also, distributary channels have well-defined sandy levees. Interdistributary areas away from wind-waves contain quiescent marshes that slowly accrete over time.
One highly distinct area on the delta that lack a definitive explanation for its origin is a large tract of high marsh cut by a tributary to OPC in the riparian forest. This marsh tract has many signs of animal activity including well-maintained marsh channels and channel-bank slides. Two monitoring stations, 6-P11 and 6-P21, were located within the area. Site 6-P21 was in a stand of Acorus calamus (sweetflag), the dominant high marsh species, while 6-P11 was located on the eastern levee of the tributary. These sites accumulated 0.21 and 0.13 g·cm-2·yr-1 of sediment, respectively. While 6-P21 received more sediment on average, 6-P11 experienced significant erosion during winter 1997 followed by a single large deposition event cause by a spring flood. The flood deposited 765 g of sediment at 6-P11. Given the high rate of overbank deposition on the tract as well as the tract’s position far upstream from the main intertidal zone, it seems very unlikely that the tract should naturally be high marsh at all without some unknown prior disturbance. Based on the processes observed on the delta, it is possible that historic animal activity or unreported human activity could be responsible for this region that lags behind surrounding areas in its evolution. Since debris jams have been seen to form shrub marshes with relict riparian tree species, they cannot be responsible for the state of the high marsh tract.

Discussion
Time series data of sedimentation and erosion on the OPC delta show that animals have a measurable impact on geomorphic and ecologic dynamics in the tidal freshwater zone. Animal-impacted sites have higher sedimentation rates and greater temporal variation in sedimentation and erosion than non-impacted sites. They also have lower or-ganic content and lower temporal variation in organic content. Field observations along with analyses of substrate characteristics such as grain sizes suggest that these differences were largely caused by a redistribution of local bed material within the disturbed area. Increased sedimenta-tion on a tile must therefore be offset by erosion elsewhere, perhaps in the newly formed channels. Even though the redistribution can be accomplished in a single season, the disturbed area does not recover to its pre-impact condition for many years. In comparison to HBW sites, animal-impacted sites at OPC had order-of-magnitude larger puls-es in sedimentation. These extreme fluctuations are thought to stem from the proximity of OPC sites to major distributary channels that are conduits for sediment, whereas the HBW sites are isolated, with the only channels present being the ones made by animals.
Both beavers and muskrats were observed to alter habitat conditions on the delta. Beavers worked for 3-6 weeks each autumn when marsh plants were dying back, whereas muskrats worked for 9-13 weeks as plants grew in spring and summer. Also, beavers channelized and devegetated more area than muskrats at any one time, but were less likely to return to the same spot year after year. Because beaver activities took place during the time when vegetation was decreasing, they exacerbated an already significant susceptibility to sedimentation and erosion. This timing led to higher sedimentation pulses at the beaver-impacted site (A6) than the muskrat-impacted site (B5) at HBW. However, muskrat activity succeeded over time in exposing the marsh to greater sedimentation and erosion on average between those same two locations.
Based on the field data, it is possible to construct a conceptual model of how animal activity impacts delta evolution. When no animals are present, spatio-temporal distributions of sedimentation and erosion in the intertidal zone of the delta are controlled by biogeomorphic boundary conditions such as plant association and elevation as well as marsh geometry in terms of distance from the subtidal front and pioneer mudflat where suspended sediment is initially entrained. As sites grow vertically, the rate of their further growth diminishes exponentially. In the face of this long-term delta evolution driven by sediment supplied from the upstream watershed, animals work hard to maintain their habitat, which was observed to be middle marsh, and preferably areas that are already somewhat disturbed. Middle marsh is preferred because it is the only intertidal habitat with plants suitable for building lodges. The geomorphically significant changes induced by animals include plant cutting, elevation reduction, and channelization. These changes cause higher sedimentation rates and reverse succession of an area from middle marsh to low marsh. It takes years for the system to aggrade back to its pre-disturbance state, partly because of the improved drainage of disturbed sites and partly due to increased winter erosion characteristic of the low marsh.
Given that animals can significantly change local habitat conditions, the question remains as to whether the ob-served animal-induced reverse succession is widespread enough to fundamentally affect geomorphic evolution at the scale of the whole delta. Presently there are too few animals to have such an impact, but historically lowland animal populations in Maryland were very large. How would such a population respond to the anthropogenically induced speed-up in delta growth? It is hypothesized that once the tidal freshwater zone evolved from open water submerged aquatic vegetation habitat to intertidal marsh, a large area of suitable habitat became available to the animals. However, as sedimentation continued, animal popu-lations must have struggled to maintain their preferred conditions. This struggle would have been more success-ful in areas that are isolated from the major distributary channels carrying anthropogenic sediment through the delta. Consideration of the delta’s biogeomorphic structure points to both HaHa Branch Wetland and the high marsh tract surrounding the small tributary in the riparian forest as examples of just such isolated areas. Both had a diverse array of intertidal and riparian habitats that could provide all the resources animals needed and both are stunted in their long-term geomorphic evolution and plant succession relative to their landscape position. Therefore, it is surmised that these sites lag the rest of the delta plain because of animal activity. Once local animal populations were reduced to a negligible size, these areas would have evolved from low and middle marsh to their current states.
Using a combination of field monitoring and landscape interpretation it is deduced that animals in the Chesapeake Bay region cannot prevent human-induced rapid delta progradation, but can alter the biogeomorphic structure of a delta. There is an important distinction between frontal progradation and vertical accretion of the delta plain that must be recognized. Animals can combat vertical accretion on the delta plain wherever there are interdistributary areas that are isolated from the primary sediment transport conduits. Such areas exist at Otter Point Creek and over the long term are two steps behind the rest of the delta plain in habitat succession and geomorphic evolution. The more isolated the site that is impacted by animal activity, the less able the system will be to restore the conditions there, as transport capacity and sediment supply are both low in these locales. Over the long term, disturbed sites provide habitat diversity, and thus animals serve an important function in the tidal freshwater river ecosystem.
Publications
- Pasternack, G. B. 1998. Physical dynamics of tidal freshwater delta evolution. Ph. D. Dissertation. The Johns Hopkins University, 227pp, 5 appendices.
- Pasternack, G. B. 2001. Animal Response to River Evolution in the Tidal Freshwater Zone. In (J. M. Dorava, D. R. Montgomery, B. B. Palcsak, and F. A. Fitzpatrick, Eds) Geomorphic Processes and Riverine Habitat. American Geophysical Union, Water Science and Application Volume 4, p. 139-157.
